 | The Nuclear pore complex (NPC) has a molecular mass of ~125 mDa in vertebrates and contains about 50 or more different proteins (reviewed in Nakielny and Dreyfuss, 1999; Lyman and Gerace, 2001)  . The NPC spans the dual membrane of the the nuclear envelope (NE) and acts as a gateway for macromolecular traffic between the cytoplasm and the nucleus.
The basic framework of the NPC consists of a central core
with a ring-spoke structure exhibiting 8 fold radial symmetry. From this central ring 50 to 100 nm fibrils extend into the nucleoplasm and the cytoplasm. The NPC is inturn anchored in the NE by the nuclear lamina, a meshwork of lamins and lamin-associated proteins that forms a 15 nm thick fibrous structure between the inner nuclear membrane and peripheral chromatin. A number of proteins called nucleoporins . The NPC spans the dual membrane of the the nuclear envelope (NE) and acts as a gateway for macromolecular traffic between the cytoplasm and the nucleus.
The basic framework of the NPC consists of a central core
with a ring-spoke structure exhibiting 8 fold radial symmetry. From this central ring 50 to 100 nm fibrils extend into the nucleoplasm and the cytoplasm. The NPC is inturn anchored in the NE by the nuclear lamina, a meshwork of lamins and lamin-associated proteins that forms a 15 nm thick fibrous structure between the inner nuclear membrane and peripheral chromatin. A number of proteins called nucleoporins
|
| have been localised to discrete regions of the NPC and are often used as
markers for this compartment, e.g. Nup153 (for further reading see: Stoffler et al., 1999). Approximately half of the nucleoporins (or Nups) contain a phenyalanine-glycine repeat motif (FG repeat), which may be diagnostic for proteins playing a role in nuclear transport.
Protein cargo identifies itself to the nucleocytplasmic transport machinery by either nuclear localisation signals (NLSs) or nuclear export signals (NESs), which can be based on protein, RNA or a consist of a composite of both (Nakielny and Dreyfuss, 1999). A number of nuclear transport receptors exist that recognise these signals, variously called karyopherins, PTACs, importins, transportins and Ran-binding proteins. | 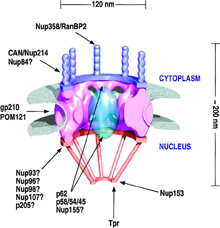 | Nuclear Pore Complex
Click to enlarge
(Nakielny and Dreyfuss, 1999) |
These receptors are generally large acidic proteins that share the ability to bind
components of the NPC and contain both an N-terminal
RanGTP-binding domain and a C-terminal cargo-binding domain. Cargo (protein, RNA or both) can bind its cognate nuclear transport receptor directly through NLS and NES signals or via adaptor proteins. In particular, proteins with simple or bipartite NLSs are imported via the b-importin receptor indirectly by association with the a-importin family of adaptors.
Nuclear transport and Ran
RanGTP Cycle
and Nuclear Import/Export
Click to enlarge
(Nakielny and Dreyfuss, 1999)
|
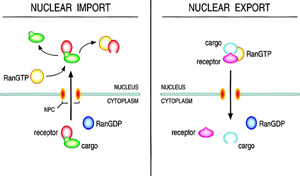
Although cargo can accumulate at the cytoplasmic face of the NPC independent of temperature, nuclear translocation of cargo is a temperature dependent phenomenon. (Nakielny and Dreyfuss, 1999). Discovery of the essential protein import factor RanGTPase, suggested that the temperature dependence of the nuclear accumulation of cargo was due to reduction in GTP hydrolysis. Consequently it has been shown that the directionality of nuclear transport, rather than actual movement through the channel of the nuclear pore, is dependent on NTP hydrolysis through Ran. To date, the mechanism that propels cargo through the NPC channel itself, remains a issue of heated debate.Ran is maintained as RanGTP in the nucleus by its association with the Ran GTP-GDP exchange factor (RanGEF or RCC1).
|
Ran is maintained as RanGTP in the nucleus by its association with the Ran GTP-GDP exchange factor (RanGEF or RCC1).
In the cytoplasm the RanGTPase activating protein (RanGAP) simulates the GTPase activity of Ran leading to the accumulation of RanGDP in the cytoplasm. In addition, RanBP1 in the cytoplasm and RNA BP1-like domains on the cytoplasmic fibrils of the NPC act as coactivators of RanGAP, which further promotes the hydrolysis of GTP by Ran. Import receptors bind their cargos in a RanGTP-indpendent manner and release their cargos in the presence of RanGTP, which accumulates in the nucleus through the action of RanGEF. In reciprocol fashion, export receptors bind their cargo in a stable manner in the presence of RanGTP but release of this cargo requires GTP hydrolysis by Ran and the accumulation of RanGDP. Thus, GTP hydrolysis by Ran is required to accumulate cargo against a concentration gradient between the nucleus and cytoplasm.
The NPC and Disease
Nuclear pore complex components are implicated in both autoimmune disease and cancer. For example, the NUP98 gene localized to chromosome band 11p15 has been found at the breakpoints of several distinct chromosomal translocations in patients with both de novo and therapy-related myelodysplastic syndromes (MDS) and acute myelogenous leukemia (AML). These translocations usually result in the generation of chimeric proteins containing sequences from NUP98. The Hox genes are common fusion partners for NUP98, including HOXD13 and HOXA9 (Nishiyama et al., 1999; Arai et al., 2000 ). In the case of the Nup98-HoxA9 fusion, expression of this fusion protein in hemopoietic stem cells can induce chronic myeloid leukemia (CML) and AML in mice (Kroon et al., 2001) . However, in a recent de novo case of acute myeloid leukemia (AML), Nup98 was disrupted by a translocation that resulted in the formation of a chimeric gene between Nup98 and a novel transcriptional regulator, NSD1 (at 5q35)(Jaju et al., 2001). The transforming ability of these nucleoporin fusion genes appears to arise form the presence of N-terminal FG dipeptides from Nup98, which have the ability to recruit CBP/p300 to, and thus activate, loci bound by the Nup98-Hox gene fusion protein (Kasper et al., 1999) .
Autoimmunity to nuclear envelope components including the NPC is important in both the diagnosis and etiology of several autoimmune diseases. Patients with primary biliary cirrhosis (PBC) frequently produce autoantibodies against the nucleoporin p62, Nup180 and gp210, an integral glycoprotein of the nuclear pore (Wilken et al., 1993; Nesher G et al., 2001) . Consequently, these antibodies have been used as diagnostic markers for PBC.
The Nuclear Envelope--From Gwen Childs, an overview of the nuclear envelope including the nuclear pore complex and nuclear/cytoplasmic trafficking. Great EM photos.Daniel Stoffler's Nuclear Pore Page
-More NPC slides
Winey Lab Pages-- NPC protocols, images and reagents
Nucleopore Antibodies -- Affinity BioReagents, Inc.
Ed Hurt's Web Pages--University of Heidelberg (overview of nuclear import/export)
Published Movies of Nuclear Pore dynamics
Daigle et al. (2001)
Nuclear pore complexes
form immobile networks and have a very low turnover in live mammalian
cells. JCB
154:74
REFERENCES
Arai Y, Kyo T, Miwa H, Arai K, Kamada N, Kita K, Ohki M. (2000)
Heterogenous fusion transcripts involving the NUP98 gene and HOXD13
gene activation in a case of acute myeloid leukemia with the
t(2;11)(q31;p15) translocation. Leukemia 14(9):1621-1629
Jaju RJ, Fidler C, Haas OA, Strickson AJ, Watkins F, Clark K, Cross NC, Cheng JF,
Aplan PD, Kearney L, Boultwood J, Wainscoat JS. (2001) A novel gene, NSD1, is fused to NUP98 in the t(5;11)(q35;p15.5) in de novo
childhood acute myeloid leukemia. Blood 98(4):1264-1267
Kasper LH, Brindle PK, Schnabel CA, Pritchard CE, Cleary ML, van Deursen JM. (1999) CREB binding protein interacts with nucleoporin-specific FG repeats that activate transcription
and mediate NUP98-HOXA9 oncogenicity.
Mol Cell Biol. 19(1):764-776.
Kroon E, Thorsteinsdottir U, Mayotte N, Nakamura T, Sauvageau G. (2001) NUP98-HOXA9 expression in hemopoietic stem cells induces chronic and acute myeloid
leukemias in mice. EMBO J. 20(3):350-361
Lyman SK, Gerace L. (2001) Nuclear pore complexes: dynamics in unexpected places. J. Cell Biol.154(1):17-20.
Nakielny S, Dreyfuss G. (1999) Transport of proteins and RNAs in and out of the nucleus. Cell 99(7):677-690
Nesher G, Margalit R, Ashkenazi YJ. (2001) Anti-nuclear envelope antibodies: Clinical associations.
Semin Arthritis Rheum. 30(5):313-320.
Nishiyama M, Arai Y, Tsunematsu Y, Kobayashi H, Asami K, Yabe M, Kato S, Oda M,
Eguchi H, Ohki M, Kaneko Y. (1999) 11p15 translocations involving the NUP98 gene in childhood
therapy-related acute myeloid leukemia/myelodysplastic syndrome. Genes Chromosomes Cancer 26(3):215-220
Spector, D.L. (2001) Nuclear domains. J. Cell Sci. 114 (16):2891-2893
Stoffler D, Fahrenkrog B, Aebi U. (1999) The nuclear pore complex: from molecular architecture to functional dynamics.
Curr Opin Cell Biol. 11(3):391-401
Wilken N, Kossner U, Senecal JL, Scheer U, Dabauvalle MC. (1993)
Nup180, a novel nuclear pore complex protein localizing to the
cytoplasmic ring and associated fibrils. J. Cell Biol.123(6 Pt 1):1345-1354